Automation from DNA and proteins: what does it have in common with electronics?
(biologist's answer to the article “You write animals incorrectly” )

Every cell has thousands of genes. It never happens that they all work at the same time. Muscle, liver and brain cells carry the same set of genes. They are so different because different genes work in them. All cells are able to turn on and off individual genes in response to various external influences. That is, the system that controls the activity of genes is such an automatic control system. Scientists want to understand how such automation works in order to be able to repair and crack it. For example, in human cells there is a system of about three hundred genes and proteins that controls cell division. When it breaks down and the cell begins to divide all the time, cancer occurs.
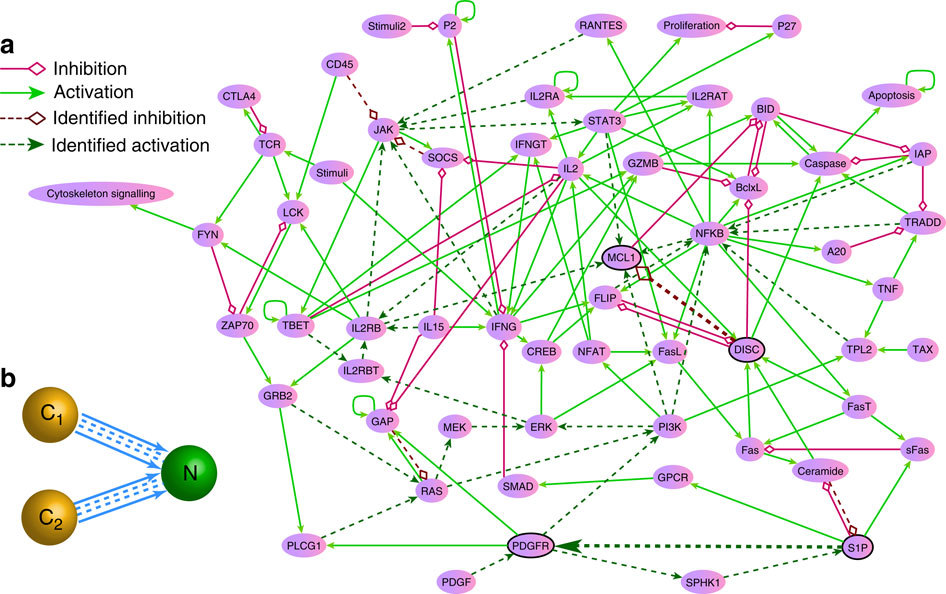
Simplified diagram of the gene network regulating the division of human cells
In the "gland" are usually used electrical impulses. Signals in biology, as a rule, are chemical, that is, changes in the concentration of some substances. There are electrical signals in the nervous system, but this is only a device for fast far transmission, like optical fiber in technology. Nerve impulse transmission from cell to cell occurs in chemical form, long-term memory and complex integration of signals in the neuron is also chemical.
')
Where there are signals, there will be pickups - signals are out of place and noises are random garbage clinging to signals. In electronics, the arrival of signals at the address is provided by wires and insulation. Leads occur due to parasitic capacitive and inductive coupling between conductors. In the cell of a bacterium its contents are more or less evenly mixed, and any chemical signal (and there are thousands of them) is available at any of its points. The cells of animals and plants are divided into compartments with different chemical composition, but there are less than a dozen types of these compartments and each has hundreds and thousands of different substances.
The arrival of the signal in the right place of the cell is due to molecular recognition by the “key-lock” principle. One protein molecule can recognize another protein, a protein can recognize a specific DNA sequence, a protein can recognize small molecules, such as sugars. This recognition is not always perfectly accurate, so molecules that are similar in shape to a regular recognition partner will create leads. In addition to interference, there are noises in electronics. They arise from the thermal motion of atoms, which introduces a fraction of chaos in the motion of electrons. In a cell, chemical noise is also associated with thermal movement. When the signal is weak, for example, ten pieces of molecules of a given type for the whole cell, these molecules will be scattered around the cell by chance, and it’s not a fact that it is uniform. As a result, their concentration in some corner of the cell will randomly fluctuate, and it will be noise.
A cell can be considered as a plant that produces all the equipment it needs and can collect from it the second such plant. The main equipment are ribosomes - CNC machines that collect all proteins. The structure of the collected protein is recorded on a tape - messenger RNA (mRNA). The ribosome moves along the mRNA and, according to its instructions, collects a chain of a new protein from individual units — amino acids. The protein chain coming out of the ribosome is folded into a compact tangle of mature protein that begins to work. Proteins can accelerate chemical reactions (enzymes), as well as engage in the transport of substances, signaling, protection, and many other things.
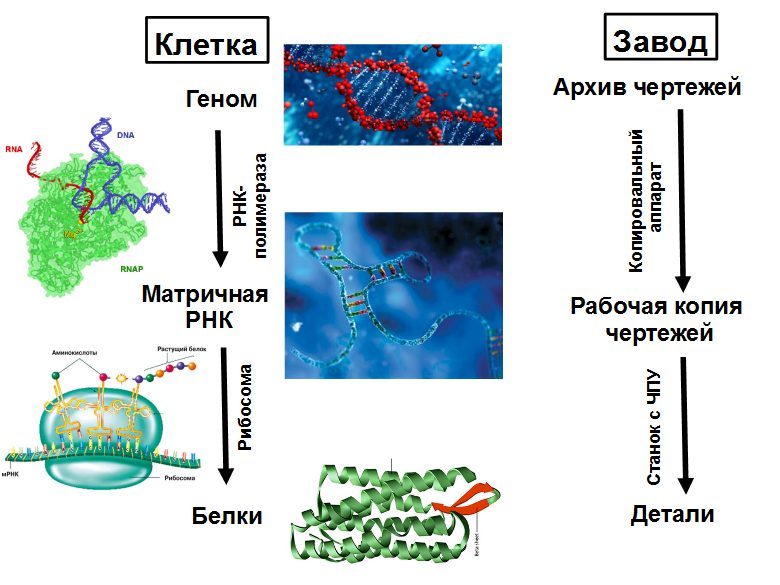
mRNAs are formed as copies of individual sections of cellular DNA. DNA can be compared with the archive of technological documentation in the office of the chief engineer, and mRNA - with copies of the drawings that are issued to the workshop. The copy machine that creates these copies is also a protein called RNA polymerase. She can sit on the DNA and start copying it not everywhere, but only on special sections of DNA — promoters that are in front of each gene or group of genes working together.
Turning on and off genes is mainly determined at the stage of RNA polymerase landing on the promoter. If RNA polymerase easily sits on the promoter of a gene, then many mRNA copies are made from it and the ribosomes produce many protein molecules encoded by this gene - it will be included. If something interferes with the landing of the RNA polymerase, then the gene will be turned off. That is, switching on and off genes occurs at the stage of interaction between the RNA polymerase and the promoter. Various other molecules can help or interfere with their binding. These other molecules, primarily the special regulatory proteins, together with the promoter and the adjacent DNA segments form a logical element that can integrate several different input signals.
Perhaps the most studied example of a gene switch is Escherichia coli lactose operon. The French Jacob and Monod were awarded the Nobel Prize in 1965 for his research. E. coli, as its name implies, lives in the intestines. This may be the intestines of a variety of animals, from bees to humans. She has to eat there what she got to the owner, so she can eat many different nutrients.

E. coli under a microscope and on a Petri dish
E. coli can grow, for example, on a nutrient medium consisting only of glucose and mineral salts and produce all the amino acids and vitamins it needs. Instead of glucose, there may be other sugars (fructose, malt sugar - maltose, milk sugar - lactose and a dozen others), as well as polysaccharides, such as starch.
For the assimilation of each of these sugars need their own enzymes. The production of these enzymes should be started only when the corresponding sugar is in the medium, otherwise the materials and energy for the production of these enzymes will be wasted. That is, the genes of these enzymes assimilation of sugars should be included only when there is this sugar. For the assimilation of each sugar, you need, as a rule, several enzymes, and not one. Their genes are located next to the DNA chain and their activity is controlled by one common regulatory DNA region at the beginning of the first gene. Such a group of jointly working and synchronously controlled genes is called an “operon”.

Lactose Operon Scheme
Lactose operon consists of three genes. The first (LacZ) encodes a transport protein that pumps lactose into the cell, and the other two (LacI and LacA) encode enzymes, the joint work of which turns lactose into a “default sugar” glucose. At the beginning of the lactose operon there is an RNA polymerase binding site (promoter) and a regulatory protein binding site (operator).
The activity of the lactose operon is controlled by two signals. The first signal is obviously lactose concentration. If there is no lactose, then its digestion enzymes are not needed. The second signal is a bit more complicated. In one experiment, Jacob and Mono raised E. coli on media containing both lactose and glucose. Under such conditions, the bacillus consumes glucose first, and its lactose operon is turned off. When glucose ends, the growth of bacteria stops for 15-20 minutes, and then continues due to lactose. During the pause, the lactose operon is activated. That is, the second regulatory input turns off the lactose operon when the cell has more available sugars than lactose (glucose uptake does not require any additional enzymes).
Further experiments showed that the lactose operon does not respond to the glucose itself. Instead, inside the cells of E. coli, the level of hunger (more precisely, the “taste” of available sugars) is encoded by a special signal substance. It is called “cyclic adenosine monophosphate”, or cAMP. While the bacillus grows on glucose, there is practically no cAMP in the cell. If there is no glucose, but there is maltose or starch (slightly more difficult to digest), a little cAMP is produced. If there is no maltose, but there is fructose or lactose, the level of cAMP in the cell will be higher. If there are no sugars, but there is lactic acid - even higher, and finally, if there is nothing tasty and you have to eat glycerin, then the level of cAMP will be the highest. So, the lactose operon is controlled by two signals: the concentration of lactose and the concentration of cAMP.
How do these signals act on gene activity? An active gene is one in which RNA polymerase produces many messenger RNAs. RNA polymerase begins to work on this operon always from landing on the same DNA segment (it is called a “promoter”). Where is the place for lactose and cAMP? RNA polymerase cannot recognize all of the hundreds of signals that regulate the activity of thousands of genes. For this, there are special regulatory proteins (they are called “transcription factors”). Two regulatory proteins are involved in controlling the lactose operon, one for each input signal: the lactose repressor and the catabolic activator.
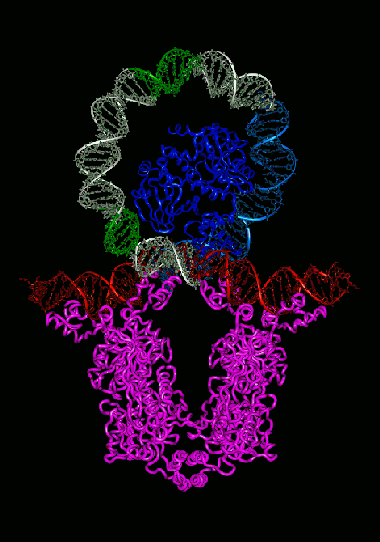
Lactose repressor (purple) on DNA
A lactose repressor is a protein that can bind to either lactose or a specific DNA sequence. There are two such sequences in the genome of E. coli, both at the beginning of the lactose operon, on both sides of the promoter. The lactose repressor molecules firmly stick together with each other in groups of four protein molecules. Binding of such a quadruple to DNA twists the DNA strand into a steep loop in which the promoter is located. RNA polymerase can not sit on such a twisted promoter and the gene is turned off. If lactose appears in the cell, it binds to the lactose repressor and it falls off from the DNA, releasing the promoter for RNA polymerase - the operon starts working. This is how the regulatory input of the operon on the concentration of lactose is arranged.
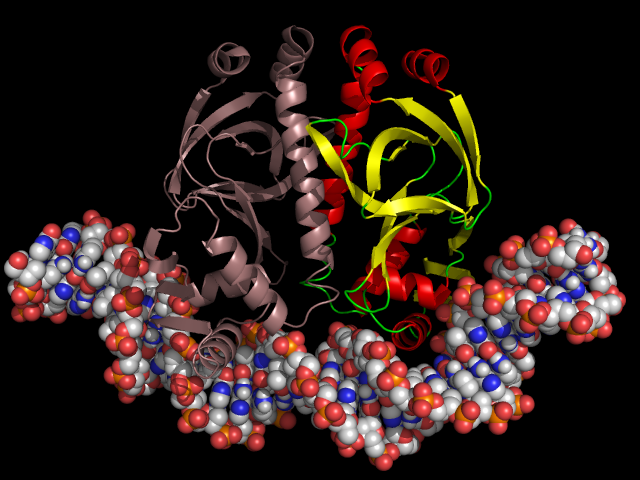
DNA catabolic activator
The catabolic activator responds in a similar way to the concentration of cAMP. True, there are a few differences in details:
- catabolic activator sits on the DNA a little to the side of the promoter and does not overlap it. On the contrary, it enhances the binding of RNA polymerase to the promoter and increases the activity of the operon.
- catabolic activator binds cAMP and DNA. Without cAMP, it cannot bind to DNA.
- a catabolic activator is needed for the full activity of the lactose operon, but even without it, the operon will still work approximately 5% of the total activity. The lactose repressor regulates the operon in the range from 0 to 100%.
- molecules of the catabolic activator are connected in pairs, not in fours. In the next post we will see what this difference leads to.
That is, we can write a truth table for the lactose operon:
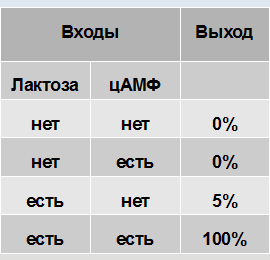
Lactose operon works almost like a logical element AND. Only 5% of the activity in the "lactose +, cAMP -" state slightly drops out of the ideal picture.
Continued with charts and matan: https://geektimes.ru/post/283686/
Every cell has thousands of genes. It never happens that they all work at the same time. Muscle, liver and brain cells carry the same set of genes. They are so different because different genes work in them. All cells are able to turn on and off individual genes in response to various external influences. That is, the system that controls the activity of genes is such an automatic control system. Scientists want to understand how such automation works in order to be able to repair and crack it. For example, in human cells there is a system of about three hundred genes and proteins that controls cell division. When it breaks down and the cell begins to divide all the time, cancer occurs.
Simplified diagram of the gene network regulating the division of human cells
How does intracellular automatics work?
In the "gland" are usually used electrical impulses. Signals in biology, as a rule, are chemical, that is, changes in the concentration of some substances. There are electrical signals in the nervous system, but this is only a device for fast far transmission, like optical fiber in technology. Nerve impulse transmission from cell to cell occurs in chemical form, long-term memory and complex integration of signals in the neuron is also chemical.
')
Where there are signals, there will be pickups - signals are out of place and noises are random garbage clinging to signals. In electronics, the arrival of signals at the address is provided by wires and insulation. Leads occur due to parasitic capacitive and inductive coupling between conductors. In the cell of a bacterium its contents are more or less evenly mixed, and any chemical signal (and there are thousands of them) is available at any of its points. The cells of animals and plants are divided into compartments with different chemical composition, but there are less than a dozen types of these compartments and each has hundreds and thousands of different substances.
The arrival of the signal in the right place of the cell is due to molecular recognition by the “key-lock” principle. One protein molecule can recognize another protein, a protein can recognize a specific DNA sequence, a protein can recognize small molecules, such as sugars. This recognition is not always perfectly accurate, so molecules that are similar in shape to a regular recognition partner will create leads. In addition to interference, there are noises in electronics. They arise from the thermal motion of atoms, which introduces a fraction of chaos in the motion of electrons. In a cell, chemical noise is also associated with thermal movement. When the signal is weak, for example, ten pieces of molecules of a given type for the whole cell, these molecules will be scattered around the cell by chance, and it’s not a fact that it is uniform. As a result, their concentration in some corner of the cell will randomly fluctuate, and it will be noise.
What exactly is regulated in the living cell?
A cell can be considered as a plant that produces all the equipment it needs and can collect from it the second such plant. The main equipment are ribosomes - CNC machines that collect all proteins. The structure of the collected protein is recorded on a tape - messenger RNA (mRNA). The ribosome moves along the mRNA and, according to its instructions, collects a chain of a new protein from individual units — amino acids. The protein chain coming out of the ribosome is folded into a compact tangle of mature protein that begins to work. Proteins can accelerate chemical reactions (enzymes), as well as engage in the transport of substances, signaling, protection, and many other things.
mRNAs are formed as copies of individual sections of cellular DNA. DNA can be compared with the archive of technological documentation in the office of the chief engineer, and mRNA - with copies of the drawings that are issued to the workshop. The copy machine that creates these copies is also a protein called RNA polymerase. She can sit on the DNA and start copying it not everywhere, but only on special sections of DNA — promoters that are in front of each gene or group of genes working together.
Turning on and off genes is mainly determined at the stage of RNA polymerase landing on the promoter. If RNA polymerase easily sits on the promoter of a gene, then many mRNA copies are made from it and the ribosomes produce many protein molecules encoded by this gene - it will be included. If something interferes with the landing of the RNA polymerase, then the gene will be turned off. That is, switching on and off genes occurs at the stage of interaction between the RNA polymerase and the promoter. Various other molecules can help or interfere with their binding. These other molecules, primarily the special regulatory proteins, together with the promoter and the adjacent DNA segments form a logical element that can integrate several different input signals.
Logic elements on DNA and proteins
Perhaps the most studied example of a gene switch is Escherichia coli lactose operon. The French Jacob and Monod were awarded the Nobel Prize in 1965 for his research. E. coli, as its name implies, lives in the intestines. This may be the intestines of a variety of animals, from bees to humans. She has to eat there what she got to the owner, so she can eat many different nutrients.
E. coli under a microscope and on a Petri dish
E. coli can grow, for example, on a nutrient medium consisting only of glucose and mineral salts and produce all the amino acids and vitamins it needs. Instead of glucose, there may be other sugars (fructose, malt sugar - maltose, milk sugar - lactose and a dozen others), as well as polysaccharides, such as starch.
For the assimilation of each of these sugars need their own enzymes. The production of these enzymes should be started only when the corresponding sugar is in the medium, otherwise the materials and energy for the production of these enzymes will be wasted. That is, the genes of these enzymes assimilation of sugars should be included only when there is this sugar. For the assimilation of each sugar, you need, as a rule, several enzymes, and not one. Their genes are located next to the DNA chain and their activity is controlled by one common regulatory DNA region at the beginning of the first gene. Such a group of jointly working and synchronously controlled genes is called an “operon”.
Lactose Operon Scheme
Lactose operon consists of three genes. The first (LacZ) encodes a transport protein that pumps lactose into the cell, and the other two (LacI and LacA) encode enzymes, the joint work of which turns lactose into a “default sugar” glucose. At the beginning of the lactose operon there is an RNA polymerase binding site (promoter) and a regulatory protein binding site (operator).
The activity of the lactose operon is controlled by two signals. The first signal is obviously lactose concentration. If there is no lactose, then its digestion enzymes are not needed. The second signal is a bit more complicated. In one experiment, Jacob and Mono raised E. coli on media containing both lactose and glucose. Under such conditions, the bacillus consumes glucose first, and its lactose operon is turned off. When glucose ends, the growth of bacteria stops for 15-20 minutes, and then continues due to lactose. During the pause, the lactose operon is activated. That is, the second regulatory input turns off the lactose operon when the cell has more available sugars than lactose (glucose uptake does not require any additional enzymes).
Further experiments showed that the lactose operon does not respond to the glucose itself. Instead, inside the cells of E. coli, the level of hunger (more precisely, the “taste” of available sugars) is encoded by a special signal substance. It is called “cyclic adenosine monophosphate”, or cAMP. While the bacillus grows on glucose, there is practically no cAMP in the cell. If there is no glucose, but there is maltose or starch (slightly more difficult to digest), a little cAMP is produced. If there is no maltose, but there is fructose or lactose, the level of cAMP in the cell will be higher. If there are no sugars, but there is lactic acid - even higher, and finally, if there is nothing tasty and you have to eat glycerin, then the level of cAMP will be the highest. So, the lactose operon is controlled by two signals: the concentration of lactose and the concentration of cAMP.
How do these signals act on gene activity? An active gene is one in which RNA polymerase produces many messenger RNAs. RNA polymerase begins to work on this operon always from landing on the same DNA segment (it is called a “promoter”). Where is the place for lactose and cAMP? RNA polymerase cannot recognize all of the hundreds of signals that regulate the activity of thousands of genes. For this, there are special regulatory proteins (they are called “transcription factors”). Two regulatory proteins are involved in controlling the lactose operon, one for each input signal: the lactose repressor and the catabolic activator.
Lactose repressor (purple) on DNA
A lactose repressor is a protein that can bind to either lactose or a specific DNA sequence. There are two such sequences in the genome of E. coli, both at the beginning of the lactose operon, on both sides of the promoter. The lactose repressor molecules firmly stick together with each other in groups of four protein molecules. Binding of such a quadruple to DNA twists the DNA strand into a steep loop in which the promoter is located. RNA polymerase can not sit on such a twisted promoter and the gene is turned off. If lactose appears in the cell, it binds to the lactose repressor and it falls off from the DNA, releasing the promoter for RNA polymerase - the operon starts working. This is how the regulatory input of the operon on the concentration of lactose is arranged.
DNA catabolic activator
The catabolic activator responds in a similar way to the concentration of cAMP. True, there are a few differences in details:
- catabolic activator sits on the DNA a little to the side of the promoter and does not overlap it. On the contrary, it enhances the binding of RNA polymerase to the promoter and increases the activity of the operon.
- catabolic activator binds cAMP and DNA. Without cAMP, it cannot bind to DNA.
- a catabolic activator is needed for the full activity of the lactose operon, but even without it, the operon will still work approximately 5% of the total activity. The lactose repressor regulates the operon in the range from 0 to 100%.
- molecules of the catabolic activator are connected in pairs, not in fours. In the next post we will see what this difference leads to.
That is, we can write a truth table for the lactose operon:
Lactose operon works almost like a logical element AND. Only 5% of the activity in the "lactose +, cAMP -" state slightly drops out of the ideal picture.
Continued with charts and matan: https://geektimes.ru/post/283686/
Source: https://habr.com/ru/post/399233/
All Articles