Simulator of the nervous system. Part 3. Associative Neuroelement

In the work on the nervous system simulator, we have so far dealt only with well-studied aspects of her work. But the complexity of modeling the nervous system and the reason for which artificial intelligence has not yet been created is the lack of a complete understanding of how the nerve cell works. Many processes occurring in the nerve cell and the nervous system as a whole are described in detail, but there is no clear algorithm for their work that could be transferred to a model or computer program.
A simple idea of the algorithm of the neuron allowed to solve this problem.
Table of contents
1. Simulator of the nervous system. Part 1. Simple adder
2. Simulator of the nervous system. Part 2. Modulated Neuroelement
3. Simulator of the nervous system. Part 3. Associative Neuroelement
4. Memory, memory consolidation and grandmother's neurons
5. Modeling emotions or electronic sense of novelty
6. Amazing cerebellum
7. Structure and starting brain settings
')
I like the analogy about drawing up a theory of how the brain works with puzzle collection from Jeff Hawkins’s book, On the Intellect. In compiling this puzzle, we lack some elements and some elements from another puzzle, but we have a large array of data about the nervous system and brain, which means we have an almost assembled puzzle, so we can roughly imagine the whole picture, and using our imagination identify the missing elements.
My goal is to create a logical model of the nervous system, we can say the creation of an outline of what is depicted on an unfinished puzzle, and it should correspond and not contradict all the existing elements of the puzzle and still be logically complete. In order to fill in the gaps some theoretical basis was created, which may seem to some disputable. But for the model at this stage, the main thing is that it allows you to emulate both internal and external observable phenomena occurring in the nervous system. Within the framework of the model, it is possible to explain many phenomena, such as memory and memory consolidation, emotions, specialization of neurons, and much more.
In the second part, we found out that there are three types of reflex activity established by academician I.P. Pavlov. If everything is very clear with the biological mechanisms of habituation and sensitization, then with the formation of conditioned reflexes it is not so simple as it seems. The fact is that the external manifestations of this mechanism are widely studied and described, there is no explanation of how this happens at the cellular level.


For example, we know that when a combination of the activity of two nerve centers with time between them forms a reflex arc. Those. subsequently, when one nerve center is activated, the excitation is transferred to the other nerve center. If we figuratively divide such a reflex arc into segments, and consider such segments as separate elements. It can be said that during the formation of a reflex arc of a conditioned reflex, switching of a directional character occurs in each segment. Each segment chooses a certain direction in which there will be a transfer of nervous excitement when it is activated. Of course, it is worth noting that this direction is not clearly defined for the segment, but may be correlated in certain values. You can even talk about strengthening the transmission in a certain direction and weakening in other directions.

When reflex is strengthened by repeated repetitions, it is possible to talk about refining and enhancing transmission in the direction for each segment. This concept leads to the conclusion that if we divide the entire cortex into similar segments, then we will observe in each a certain orientation in the direction with different accuracy and strength. Each segment will be dried up by a part of some kind of reflex arc of conditioned or unconditioned reflex. Presumably this orientation in the learning process may be refined or changed.

If you turn to the neural paradigm, then it does not provide for directional orientation. We have a membrane and dendrites that receive signals and an axon, through which the signal is transmitted further to other cells after space-time summation, that is, the signal is transmitted in one direction along the axon to its terminations. But at the same time, we still observe the formation of a directional spread of excitation in the brain, with the formation of conditioned reflexes.
Neuron Paradigm
This idea of a neuron is more likely formulated by cybernetics than by neuroscientists, but also among physiologists it is input. Everything is somewhat more complicated. First, neurons are afferent, i.e. their axon brings neural impulse to the cell body and naturally further it spreads through the dendrites. Secondly, in addition to the dendritic synapses, there are dendrodendritic ones. Thirdly, existing neurons and without axons. Most likely the neuron works in all directions, its membrane is the receiver, including the membrane on the dendrites. Dendrites, like roots, grow in different directions in search of other neurons, at their tips there are transmitting synapses. If the neuron is activated, and it does not matter in which part of the membrane, then all dendrite and axon synapses will be activated. But the amount of mediator secreted will be different in different synapses, sometimes even absent altogether.
If we consider not a separate cell as a functional unit of directional switching, but a small area of cells, then we can see that the cells and their processes are very closely intertwined, and in different directions. This provides an element of directional communication with multiple inputs and outputs in various directions.

The shape of the neuron is due to evolutionary changes. The cell form was formed in the nervous systems in which only the simplest functional of the nervous activity was carried out. When the development of life on Earth required the formation of sensible reflexes to the set of functions of the nervous system, evolution did not follow the path of cell restructuring, but an increase in their number and the tight interweaving of their processes.
Thus, the property of directional switching is distributed in groups of neurons, in changing the strength of their synapses. An associative neuroelement is a functional unit in modeling and therefore an analogue in biology for it is a group of neurons for which the phenomenon of directional switching will be expressed.
We found out that the direction of the spread of excitation is important for us, but how does the determination of this direction occur for each functional element. It is known that arousal tends to spread to another source of arousal, and a stronger and larger center of arousal attracts weaker ones (the conclusion of Pavlova I.P.). Those. if a functional element gets excited, then somehow it must determine the direction that will subsequently be formed and remain in its structure.
In my work on modeling, I was repelled by the idea of electromagnetic interaction of nerve cells , and this idea gave answers to many mysteries about the brain, gave a theory and a model explaining many aspects of the nervous system.
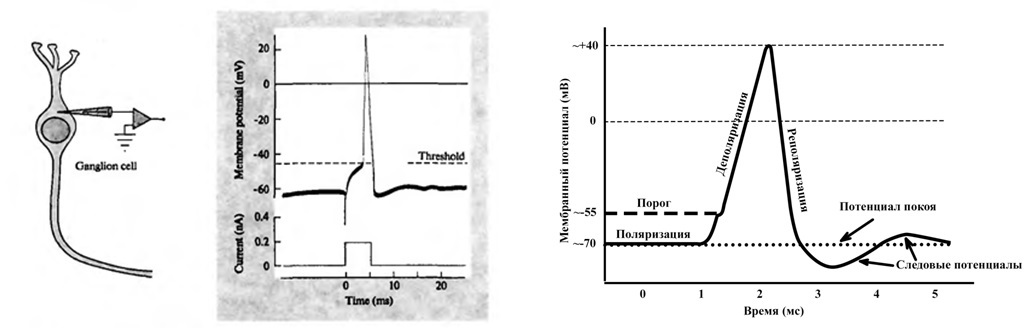
The nerve impulse throughout the nervous system has the same shape, and by analogy with it, the associative neuroelement has a charge property characterizing the change in the total charge on the membrane surface of a functional unit. Those. given a certain law of change of some characteristics called charge.
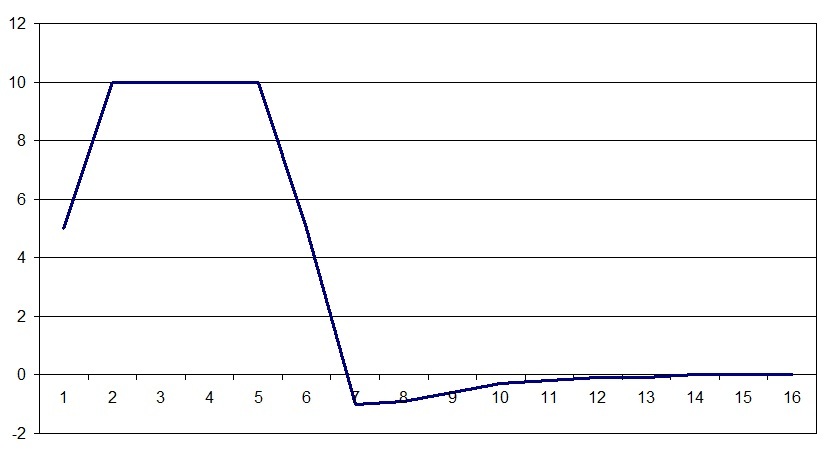
So the law is set in the program, the horizontal scale is the time in hundredths of a second, the vertical charge is in relative units. It is somewhat different from the spike graph in that part of the maximum is longer in time. This is due to the fact that the spike values are determined at one point of the nervous tissue during the passage of excitation, and the charge graph is a reflection of the charge over all the surface of a cell or group of cells. The state of rest of the nervous tissue is also taken as zero on the charge scale. It should be noted that the law of change in charge also reflects the trace potential, which was previously considered a consequence of some oscillation or equalization of charges separated by a membrane, but for the model this charge behavior turned out to be very important.

The figure above shows a diagram of an associative neuroelement. Signals from synapses of direct action (X1, X2, X3 ... Xn) are fed to the adder (a). And if the resulting amount exceeds a certain threshold (b), then the neuroelement will be activated. When a neuroelement is activated, its charge will begin to change in accordance with the established law (c). Information about these changes and the location of the item itself will be available to the entire system. Then, at a certain point in time, the mechanism for determining the vector of the preferred direction of propagation of the excitation (g) starts. This occurs by obtaining a certain average charge position of all active neuroelements, i.e. center of mass of charges, characterized by a point in space. This point is called the pattern point, because for each combination of active cells and the state of their charges at the calculated time point for each neuroelement, the position of this point will be different. Simply put, the charges of neuroelements affect the determination of the direction vector of the preferred propagation of excitation, the positive charge attracts the excitation, the negative charge repels.
To determine the vector of the preferred propagation of excitation, the following rule was chosen:

where r is the vector whose beginning is in the center of the neuroelement for which the vector is determined, and the end is in the center of the nth neuroelement.
The rule and the law of change of charge was chosen empirically, so as to simulate the formation of conditioned reflexes. More in the article .

After obtaining the vector of the preferred direction of propagation of excitation (T), the synapse force is calculated (Y1, Y2, Y3 ... Yn). Each synapse is characterized by a synapse vector (S), the beginning of which lies in the center of the neuroelement and the end is connected with the center of the target neuroelement to which the signal is transmitted. The main parameter of the synapse is its strength F, the value of the force is limited within certain limits, for example, an incentive synapse can have values from 0 to 10.
Imagine that vector T forms around itself a certain cone whose top is in the center of the neuroelement, and the base plane is perpendicular to vector T, if the synapse vector falls into the area bounded by this cone, then the value of the synapse force will be increased by a certain value. And accordingly, if the synapse vector is outside the cone area, then the synapse force decreases, but the force value does not go beyond the set maximum and minimum.
The area of the cone around the vector T is characterized by the angle at the vertex of this cone; this angle is called the focus. The smaller the focus, the more precisely the direction of excitation transfer in the neurocell will be determined. As mentioned earlier, when the body repeats the same conditioned reflex, it is refined. Therefore, the following method was chosen for the model to change the focus, when calculating the vector T, it is compared with its previous value, and if the vector changes insignificantly, the focus decreases by a certain value, but if the vector was changed greatly, then the focus returns to its maximum value. This leads to a gradual decrease in focus with repeated repetition of the same conditions.
Here is a very important point, this is how much the strength of the synapses will change with each activation. This is determined by the parameter neuroplasticity P.
The formula for the new synapse strength value will be:
Fnew = Fold + I × P × (Fmax - Fmin);
Fmin ≥ Fnew ≥ Fmax;
where P is neuroplasticity (0 ≥ P ≥ 1);
I - the parameter that determines whether the synapse vector is within the region of increasing synapse strength (I = 1) or in the region of decreasing synapse strength (I = -1);
Fold - the previous value of the synapse strength;
Fmin - the minimum value of the synapse strength;
Fmax - the maximum value of the synapse strength.
Neuroplasticity in biology describes how a neuron is pliable to changes in its structure under the influence of external conditions. For different areas of the brain is characterized by its degree of plasticity, as it may vary depending on some factors.

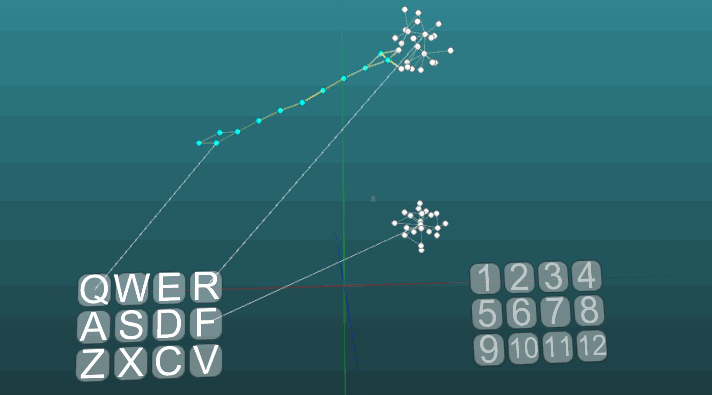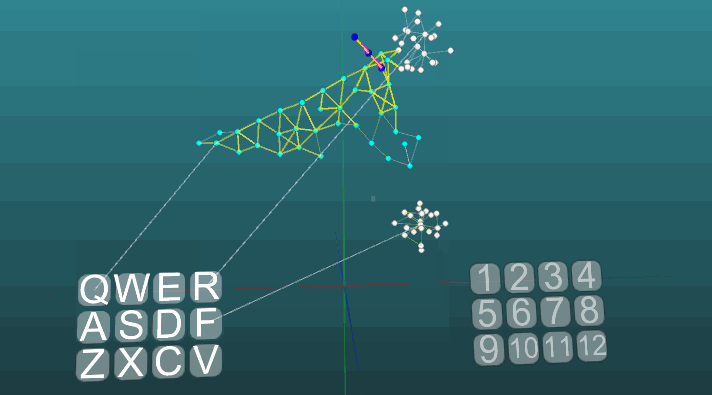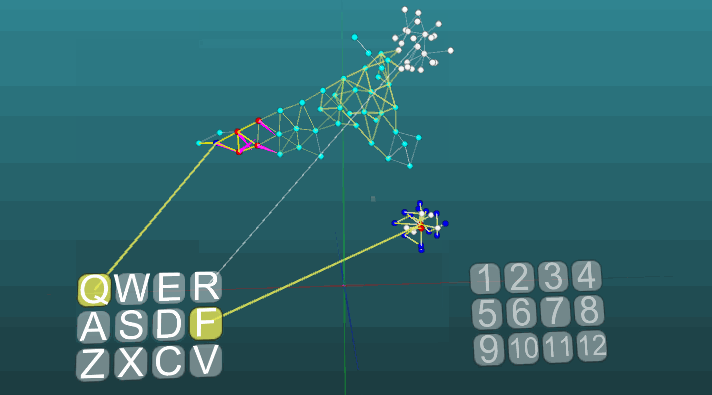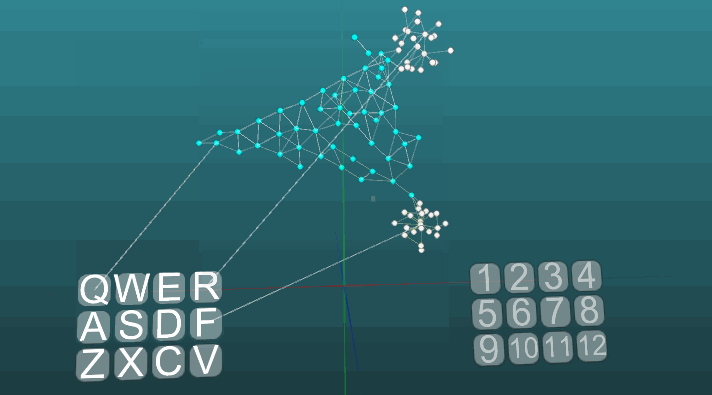
This example allows us to understand how conditioned reflexes are formed on the basis of associative neuroelements. White neuroelements form a reflex arc of an unconditioned reflex with the heading “R” and the answer “1”. These neuroelements do not change the values of the forces of their synapses. Blue neuroelements do not initially participate in any reflex acts, they seem to fill the rest of the space of the nervous system, and they are randomly interconnected through synapses. Therefore, if we activate one such neuroelement associated with the “Q” receptor, then there will be a certain focus of excitation having a random distribution pattern and going into self-centered self after some time it will go out without creating any answer. If we combine the unconditioned reflex with the heading “R” and the activation of the “Q” receptor in approximately the same time interval, a reflex arc of the conditioned reflex will be formed. And already the activation of just the “Q” receptor will lead to the answer “1”.
For clarity and optimization of the model, a dynamic creation of neuroelements was used, which emulates the occupied space of the nervous system by elements that are connected by accident. Some growth of new neurons or new connections is not modeled here, all changes occur only in the strength of synapses, just the neuroelements not previously involved in any kind of reflex act are not shown.

The following example shows how excitations behave when different centers are activated under equal conditions and with absolute plasticity (P = 1).
The change in the direction of propagation of excitation under the influence of two excitation centers, when plasticity is absolute (P = 1):
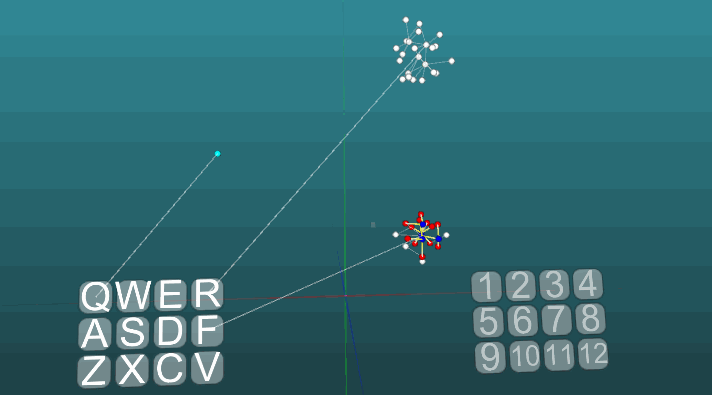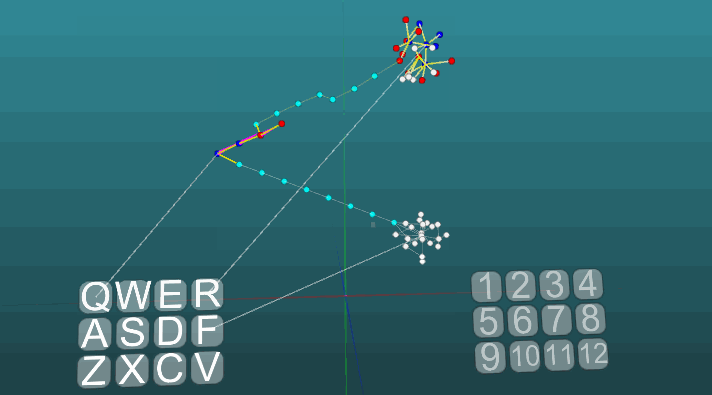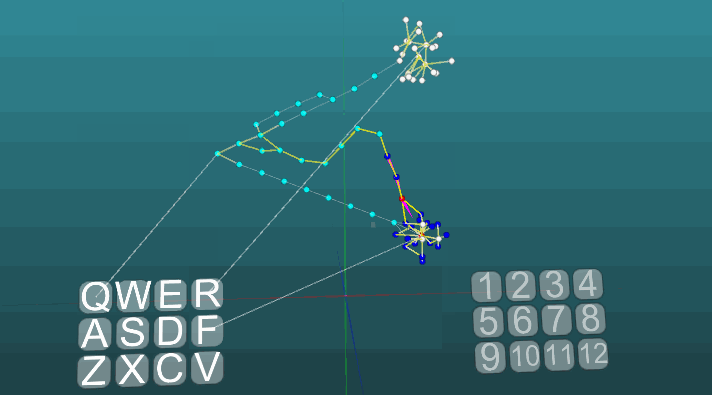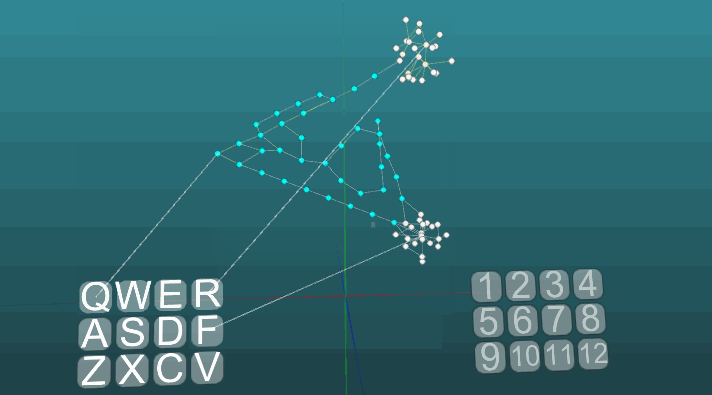
And with low plasticity (P = 0.1):
At this point we finished to consider the basics of the model of the nervous system. In the next part, we will look at applied things, how to use all this to imitate memory, emotions, and the specialization of neurons.
Source: https://habr.com/ru/post/397441/
All Articles