Nuclear thinking eukaryotes (scientific fantasy)
Introduction
The eukaryotic genomes are characterized by the presence of a large number of introns, micro- and minisatellites, dispersed elements, "relic retroviruses" and other nucleotide sequences, the total length of which exceeds the length of the gene exons that determine the primary structure of proteins by more than an order of magnitude. It is usually considered that excess DNA is a relic of evolution and only clogs the cell nucleus. At best, it plays the role of a radioprotector protecting genes from damage / 1 /.
Such explanations are sufficiently plausible, but not convincing enough. It is difficult to convince ourselves that our genome is a dustbin, full of evolutionary debris or even selfish nucleotide sequences harmful to the body. The possible radioprotective role of this “garbage” is too weak a consolation.
Especially contrasting redundancy in the human genome manifested itself after the completion of its sequencing. It turned out that the haploid set of chromosomes contains less than 30 thousand genes, and about 100 thousand nucleotides per gene. This means that less than 2% of chromosomal DNA is involved in protein coding.
')
The paradox of redundancy of the genome and the incomprehensibility of the molecular basis of the human mind are the most intriguing mysteries of modern biology. And what if these two puzzles have a common clue?
Computer analogies
Attempts to understand the principles of the human brain, as a rule, are based on the search for analogies between the structure of the central nervous system and the computer system. At the same time, nerves play the role of conductors of electrical impulses, and brain cells are equated with cells for storing and processing digital information. But the basis of the brain's ability to perceive and process information may be based on other principles. For example, analog computers are almost forgotten, which allowed to model some technological and even biological processes with the help of resistances, capacitors, transistors and other elements of electrical circuits. Other ways of processing both digital and analog information are possible.
In recent years, special attention has been paid to the theoretical and experimental elaboration of the idea of a quantum computer that operates not with bits, but with qubits, i.e. with quantum bits, whose carriers are photons, electrons, atoms, or excited molecules. The information contained in qubits is processed in parallel, which requires the use of special algorithms, but the implementation of this approach will make it possible to create microcomputer (molecular) size supercomputers capable of performing the most complicated calculations almost instantly / 2 /.
Of course, the brain is unlikely to contain quantum logical elements of the atomic or photon type, but the basis of thinking may lie in the interaction between complex molecules, leading to a change in their chemical structure or conformation. Molecular logical operations should have much in common with quantum computing and with the peculiarities of quantum computers, which include parallelism of information processing, nanometer dimensions of logical elements, as well as high efficiency calculations. For some algorithms, the efficiency of quantum computers exceeds the performance of modern digital supercomputers by orders of magnitude, and this can be achieved with only a few dozen or hundreds of logic elements / 3 /. Consequently, the efficiency of information processing is not related to the size of the computing device.
Central Bioprocessor
At the present level of development of molecular biology, it is very difficult to imagine that some “clever molecules”, which store and process incoming information, could remain unnoticed in the human brain. If thinking is really based on molecular elements, then their chemical nature is not so difficult to predict, since the phrase “informational molecules” has long been strongly associated with DNA and RNA. It is nucleic acids that are able to interact with each other with high specificity, replicate, transcribe, acquire a complex secondary and tertiary structure, change the conformation, be fragmented and even be edited.
Conditions for the interaction of information molecules exist only in the cell nucleus. It concentrates all the chromosomal DNA and a significant part of the RNA. This means that the core is able to serve as a central bioprocessor.
The cytoplasm is separated from the nucleus by a membrane that transports RNA molecules involved in gene expression with high selectivity, i.e. the very molecules whose purpose is quite understandable. Consequently, the cytoplasm does not contain (or almost does not contain) the information molecules encoded by the redundant part of the genome, and can claim only the role of a peripheral device connected to a bioprocessor.
If this assumption is true, then a multicellular eukaryotic organism should be considered a multiprocessor device consisting of a set of autonomous nuclear bioprocessors. Information from the nucleus to the cytoplasm is transmitted in the form of mRNA that encodes proteins, but for the intercellular (inter-nuclear) exchange of information, some secondary chemical or physical signals should be used, since cell membranes limiting the cytoplasm are usually impermeable to nucleic acids.
Molecular Biocomputer
The idea of using nucleic acids as molecular elements of computing devices is not new / 4, 5 /. There is even a wave genome theory that ascribes the ability of DNA to perceive, process and emit information in the form of modulated electromagnetic radiation / 6 /. Unfortunately, such ideas and theories do not allow us to understand the essence of elementary logical operations on which nuclear thinking can be based. In addition, these operations may not be elementary at all, since the algorithms of the simplest quantum computer work are hardly perceived by few specialists, and the algorithms of the molecular biocomputer may turn out to be several orders of magnitude more complicated.
The interaction of information molecules with each other occurs in parallel, and it is almost impossible to describe the sequence of events due to the absence of the sequence itself. Imagine that you did not read this text by words, but took it in its entirety. Then, without making any individual conclusions, change your view of the world around you. Perhaps this is how a nuclear bioprocessor works, in which the result of information processing is a change in the state of the system of information molecules.
Structural Linguistics
Sequences of nucleotides that provide information functions must obey some “grammatical” rules. Apparently, there are also rules of “spelling”, determined by the laws of formation of the secondary and tertiary structure of nucleic acids, as well as by the ratio of the structure of molecules to their informational properties. The study of such rules can be the main task of structural linguistics / 7 /.
Eukaryotic genomes contain up to 50% of satellite DNA, consisting of blocks of tandemly repeated nucleotide sequences. There are microsatellite (up to 4 bp in repetition) and minisatellite (more than 4 bp) DNA. Blocks of complementary nucleotide repeats can interact with each other and participate in the intermolecular interactions necessary for bioprocessor processing of information and / or in the regulation of gene expression / 8 /.
Another type of repetition includes sequences that are subdivided into SINE (short) and LINE (long) dispersed elements. The length of the SINE elements is within 400 bp, while for LINE elements it reaches 7,000 bp. The human genome contains about 1 million Alu-repeats related to SINE-elements, and more than 100 thousand long MaLR-repeats / 1 /.
The common features of dispersed elements are their transcribable and transposable ability. These features can be determined by the participation of such nucleic acids in the work of a nuclear bioprocessor. Transcribed, for example, may be associated with the need to translate double-stranded DNA into single-stranded RNA capable of intermolecular interactions. Transposition is able to move dispersed elements and change their interposition, leading to a functional adaptation of the structure of bioprocessor DNA.
The most mysterious feature of the structure of eukaryotic genes is their mosaic intron-exon structure. Genes without intron insertions are a rare exception in eukaryotes. Exons, as a rule, encode the transmitted sequences, but the purpose of introns is completely incomprehensible. The most attractive hypothesis explains the mosaicism of genes by the existence of a complex system of regulation of their expression at the level of splicing, and the interaction of introns with yet unexplored regulatory sequences / 9 / takes part in such regulation.
The average length of exons is 137 bp, whereas the length of introns may exceed 100,000 bp. /one/. It is believed that as a result of splicing, introns are removed from the mRNA, but in most cases it is more correct to consider this process as the controlled removal of exons from a mosaic molecule, the main part of which is represented by introns.
After splicing, the mRNA goes into the cytoplasm, while numerous loop-like molecules of intron RNA remain in the nucleus. Such molecules are able to carry information about the amount and nature of RNA entering the cytoplasm, and / or directly regulate the maturation of pre-mRNA through a complex network of specific intermolecular interactions. The probability of the existence of a global multimolecular multifunctional bioprocessor network, in which introns function as gene expression regulators, cannot be ruled out; moreover, this regulation is carried out both through splicing and through numerous enhancers (transcription inhibitors).
The RNA synthesized in the nucleus is combined into heterogeneous nuclear ribonucleoprotein complexes (gnRNP) associated with the components of the splicing apparatus and the nuclear matrix. Diffuse distribution in the cytoplasm is characteristic of the gnRNP complexes, but some of them can concentrate in the vicinity of the speckles (see below) and even move into the cytoplasm and return to the nucleus again.
Some facts indicate that the transfer of RNA from a gene to a nuclear membrane is not a consequence of simple diffusion. And the location of the genes, in accordance with the nuclear fixation model (gene gating model), can be functionally connected with certain sections (and pores) of the nuclear membrane / 1 /.
Core architectonics
The nucleus is surrounded by a complex shell, to the inner side of which the electron-dense layer of a nuclear lamina is attached. Nucleoplasm permeated with protein scaffold (skeleton or nuclear matrix) contains chromatin - ordered structures containing DNA, as well as histones and nonhistone proteins.
Depending on the degree of DNA compaction, euchromatin, in which the major part of the genes is expressed, and heterochromatin, which has a reduced transcriptional activity, are distinguished. Heterochromatin, in turn, is subdivided into constitutive, which often consists of repetitive DNA sequences, and optional, formed at certain stages of the organism's life cycle and usually present only in one chromosome of a pair of homologues. Structurally different regions of chromatin can be flanked by insulators — border nucleotide sequences involved in the activation of gene transcription.
Three levels of chromatin structural organization are distinguished in eukaryotes: 1) nucleosomal fibril; 2) solenoid (nucleomer); 3) a loop-domain structure, including chromomeres / 1 /. Chromomeres are formed as a result of the specific interaction of the nuclear matrix with the MAR / SAR-DNA sequences (Matrix Associated Region / Scaffold Associated Region) / 8 /. Topological domains secreted by MAR / SAR sequences may change in the process of individual development of the organism, which is accompanied by a change in the expression pattern of large blocks of genes in differentiating cells. In addition, these sequences are involved in the formation of the axis of chromatids, which together with chromatin loops form the body of the metaphase chromosome / 1 /.
The distribution of hetero- and euchromatin sites in interphase chromosomes is ordered and, apparently, can serve as a species trait. The telomeric regions of chromosomes are located near the shell, while actively expressed genes are localized mainly in the inner parts of interphase nuclei. It is shown that individual chromosomes in the nucleus do not mix and occupy discrete zones, on the periphery of which there are components of the splicing apparatus and tracks of the synthesized RNA / 1 /.
The largest non-chromosomal compartment of the nucleus is the nucleolus - the nucleoprotein complex, in which there are three main highly ordered zones: the fibrillar center, the surrounding fibrillar zone and the outer granular region. It is believed that the observed changes in the structure of the nucleolus are determined by its function of the intranuclear micro compartment of ribosome biogenesis. Recently, data have emerged indicating the involvement of the nucleolus in the regulation of the cell cycle / 1 /.
Morphologically, nucleoplasms distinguish coils of interlaced strands, which are often associated with the periphery of the nucleolus (folded bodies), dense fibrillar rings (nuclear bodies PML), nuclear domains WT1 and Barr bodies (compact heterochromatin inactive X chromosome). Perichromatin fibrils - ribonucleoprotein complexes with a diameter of 3 ... 20 nm are located at the boundaries of the condensed chromatin domains. The components of the splicing apparatus that removes introns from mRNA precursors are also detected here. In addition, RNA synthesis is closely associated with discrete inter-chromatin granules, called speckles, each core containing from 20 to 50 speckles / 1 /.
Unfortunately, the study of nuclear architectonics is associated with great methodological difficulties, but the accumulated information is quite enough to state the extremely high complexity of the structural organization of the cell nucleus. This cannot serve as evidence of the existence of nuclear bioprocessors, in which information flows are processed by molecular logic elements, but it allows not to reject such a possibility a priori.
Reasonable variability
If a molecular system with elements of consciousness arose in the process of evolution, then its main task should be self-improvement. This could lead to the emergence of mechanisms for editing nucleotide sequences that alter the coding potential of mRNA or specifically target the bases of genomic DNA.
In recent years, several RNA editing mechanisms have been discovered at the post-transcriptional level. It has been shown, for example, that in mitochondria of slugs, cytosine residues are inserted into multiple sections of RNA. In the mitochondria of trypanosomes, complex nucleoprotein complexes - editosomes / 1 / probably take part in the editing of RNA.
In the mitochondrial and nuclear transcripts of vertebrate animals, polyadenylation of mRNA results in meaningless codons, leading to premature termination of translation and the synthesis of new polypeptide chains. In the mitochondria of higher plants, as a result of enzymatic deamination, some C residues are replaced by U. A similar mRNA editing mechanism was also found in chloroplasts / 1 /.
The study of post-transcriptional editing of mRNA is just beginning, but it is likely that a similar phenomenon, which occurs in prokaryotic microorganisms, is a rule for eukaryotes, and not an exception. Much more problematic is the question of whether eukaryotes have molecular mechanisms for the correction of genomic DNA that can influence the evolution of animals and plants.
The existence of directed (adaptive) genetic variability is denied by modern science, therefore, the detection of a statistically significant increase in the incidence of Lac + E. coli revertants on a medium with lactose / 9 / almost did not become a “heresy” capable of livening up the old neo-Marcist dispute with neo-Darwinian. But everything ended successfully with the emergence of several hypotheses explaining this phenomenon on the basis of the postulate of nondirectionality of variability, proved back in the middle of the last century with the help of experiments all with the same E. coli.
The non-directionality of variability, which is one of the main axioms of the synthetic theory of evolution, has many opponents / 12, 13 /. Their objections do not contradict the evidence obtained in experiments with prokaryotes, since they are based mainly on the laws of eukaryotic evolution that are inexplicable from the point of view of neo-Darwinism. Unfortunately, the mere statement of such facts is not enough to prove the rationality of the genetic variability of eukaryotes. Modern science requires that such evidence be based on the achievements of molecular genetics. For this it is necessary to consider the mechanisms of DNA modification that can be used by eukaryotes for self-improvement.
The simplest variant of genetic adaptation is reversible duplication of genes, characteristic of both bacteria and eukaryotes. The duplication of genes determining the synthesis of ribosomal RNA is controlled at the molecular level, but this type of adaptive variability can be ranked among the usual mechanisms of regulation of gene expression.
More complex adaptive modifications of the genome may be associated with DNA methylation. The content of methylcytosine in eukaryotic DNA reaches 7% / 11 /. Eukaryotic (cytosine-5) -DNA-methyltransferase acts mainly on double-stranded DNA, one of the strands of which already contains the methylated CpG dinucleotide. This specificity of the enzyme ensures the heritability of traits determined by the replacement of cytosine with methyl cytosine. DNA methylation is one of the main factors for the fine regulation of gene expression and is likely to be able to participate in epigenetic adaptation, accelerating the evolution of eukaryotes. An indirect indication of the participation of methylation not only in microevolution, but also in the formation of new species, may be obscenely high similarity of the sequenced human and chimpanzee genes.
So far, there is no evidence of the involvement of DNA methylation in the adaptive variability of eukaryotes, but its participation in the epigenetic inheritance of some signs no longer causes any special doubts. Methylation may well underlie some manifestations of the adaptive evolution of eukaryotes. The only question is how much such an adaptation is manageable, i.e. whether the genome is capable of modifying itself, and not by trial and error, but quite purposefully.
One of the already known mechanisms for the targeted adaptation of eukaryotic cells is chromatin diminution, i.e. programmed loss of a significant part of chromosomal DNA during the formation of certain multicellular organisms / 14 /. Record on the proportion of the genome to be eliminated while Cyclops kolensis belongs to Cyclops. Only 6% of the stem cell genome is preserved in the somatic cell nuclei of this cyclop / 15 /. Less pronounced genetic rearrangements and deletions of somatic cell chromosome sections are the rule rather than the exception. The main feature of such genetic modifications is their strict genetic determinism.
Programmed rearrangements of somatic cell chromosomes fall out of the view of evolutionists, since such modifications of the genome are not inherited. But if programmed chromatin diminutions and other chromosomal rearrangements are associated with quite meaningful actions of the nuclear bioprocessor, then the possibility of a reasonable genetic adaptation of eukaryotes, which determines the high rate of their evolution, cannot be excluded.
Instead of conclusion
Cell nucleus
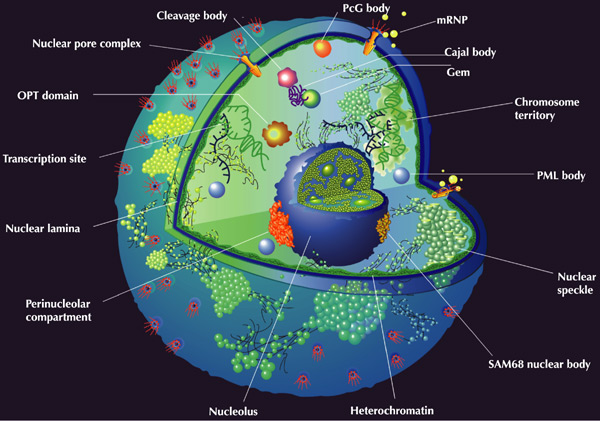
embor.embopress.org/content/4/1/18
Trash DNA is involved in brain development:
https://news.mail.ru/society/32293040//16 /
BIBLIOGRAPHY
1. Patrushev L.I. Gene expression. - M., "Science", 2000 - 527 p.
2. Valiev K.A., Kokin F.F. From the results of the 20th century: From quantum to quantum computers // aakokin.chat.ru/xx.htm
3. Valiev K.A. Quantum informatics: computers, communications and cryptography // Bulletin of the Russian Academy of Sciences. - 2000 - T. 70, No. 8. - p. 688-685.
4. Garyaev P.P., Macedonian S.N., Leonova E.A. Biocomputer on genetic molecules as a reality // Information technologies. - 1997 - № 5.
5. Luchinin V.V., Maltsev P.P. Biomolecules as the basis of future information systems // Information technologies. - 1997 - № 5.
6. Garyaev P.P. Wave genetic code. - M., 2000
7. Strugatsky A.N., Strugatsky B.N. Attempt to escape // In Sat: Favorites. - M., “Moscow Worker”, 1989 - p. 289-379.
8. Davidson EH, Britt RJ Regulation of gene expression: possible role of repetitive sequvences // Sience. - 1979. - Vol.204. - P.1052-1059.
9. Glazkov M.V. Hinge-domain organization of genes in eukaryotic chromosomes // Molecules. biology. - 1995. - T.29. - with. 965-982.
10. Miller JH Spontaneous mutators in bacteria: insight into pathways of mutagenesis and repair // Annu. Rev. Microbiol. - 1996. - Vol.50. - P.625-643.
11. Lewin B. Genes / Trans. from English - M., “Peace”. - 1987. - 544 s. (pp.295-296).
12. Lyubishchev A.A. Problems form systematics of the evolution of organisms: Sat. articles. - M., "Science". - 1982. - 278 s.
13. Berg L.S. Nomogenesis, or evolution on the basis of regularities // In Proc .: Berg LS Works on the theory of evolution. - 1977. - L., “Science”. - P.95-311.
14. Akifev A.P. Chromatin diminution - a phenomenon intended to study a number of key issues of genetics // Vavilovsky journal of genetics and selection. - 1998. - №6 (http://www.bionet.nsc.ru/vogis/vestnik.php?f=1998&p=6_5)
15. Grishanin A.K., Khudoliy G.A., Shayhaev G.O. and others. Genetics. 1996. T. 32. P. 492.
16. Ultraconserved enhancers are required for normal development / DE Dickel, et al. // Cell. - 2018. - in press.
Source: https://habr.com/ru/post/371019/
All Articles